

RESUMEN V.1
INTRODUCCIÓN
NATURALEZA BIOLÓGICA DEL LENGUAJE HUMANO
Argumentos favorables
Adquisición espontáneo y temprano
Pobreza del estímulo
Uno de los argumentos más utilizados es el de la pobreza del estímulo. Parece que el niño aprende mucho más, y más deprisa, de lo que cabría esperar de los estímulos que lo rodean. Los niños están inmersos en un mar de emisiones inconexas, fragmentarias y asistemáticas y, sin embargo, en muy poco tiempo son capaces de extraer las complejas reglas morfosintácticas que subyacen a los estímulos, incluso hasta el punto de aplicarlas inicialmente a los casos que no corresponden («rompido» por roto, «sabo» por sé).
Pongamos un ejemplo para entenderlo mejor: el sistema de reglas que rige la referencia anafórica o pronominal, que toda lengua posee como parte de su núcleo duro. En la frase siguiente: ¿Recuerdas que Miguel y sus hermanos fueron al colegio por el atajo y les salió un perro enorme?, pues es lo que le ha pasado hoy a Ana. Les se refiere de forma compacta a Miguel y sus hermanos y la partícula lo «captura» el significado global del episodio, es decir el hecho de ir al colegio a través de un atajo y de que salga un perro enorme. Todas las lenguas cuentan con recursos anafóricos como éstos, gracias a los cuales los significados se desplazan de unas partes a otras de la oración. Son relaciones de correferencia, en las que ciertos elementos inicialmente vacíos (anáforas o pronombres) se cargan de contenido al compartir referencias con otros elementos del discurso/texto. Sería inimaginable una lengua en la que hubiera que repetirlo todo cada vez. Cómo se transfieren las referencias de unas partes del discurso a otras es algo que ningún padre o madre enseña a su hijo, porque ellos tampoco reparan en ello. Este tipo de reglas son más complejas de lo que puede parecer a primera vista. Dependiendo de la estructura sintáctica, la referencia está permitida en algunos casos y bloqueada en otros: Juan sabe que él está muy enfermo. El sabe que Juan está muy enfermo. En la primera frase, la correferencia es libre y tanto Juan como él pueden referirse a la misma persona (aunque no obligatoriamente). En la segunda frase, la correferencia está bloqueada, y Juan y él son necesariamente personas distintas. No es una simple cuestión de orden; aquí la correferencia es libre aunque el pronombre va delante: Muy cerca de él, mi amigo Juan vio una serpiente. Este y muchos otros aspectos nucleares del lenguaje no son enseñados explícitamente a los niños, sino que éstos los infieren por sí mismos. Por otra parte, lo que padres y adultos enseñan del lenguaje tampoco obedece a una pauta sistemática o a algún plan didáctico que gradúe cuidadosamente la dificultad.
Todas las comunidades tienen lenguaje
Una vez constituido el Homo sapiens, todas las culturas de las que hemos tenido noticia disponían de una lengua. La historia de las civilizaciones es rica en influencias mutuas, en invasiones, en relaciones de dominancia cultural y lingüística, pero todos los pueblos, invasores o sometidos, han contado originalmente con su propia lengua.
Principios universales en todas las lenguas
Todas las lenguas del mundo, sin excepción, obedecen a un conjunto de principios universales. Y esto es así aunque muchas comunidades hayan pasado su existencia sin contacto con el resto del planeta (tribus amazónicas, etc.). Todas las lenguas están construidas sobre la base de la dualidad de estructura o doble articulación: fonemas-palabras y palabras-oraciones. En todas el signo lingüístico es simbólico y arbitrario, sin semejanza física con el referente. Y todas ellas ofrecen su extraordinaria potencialidad creadora, la explosión productiva que caracteriza al lenguaje humano. En todas las lenguas, la unidad básica del significado es la oración, formada por un sujeto y un predicado. Todas tienen sustantivos, verbos, adjetivos y pronombres, es decir, las partes principales de la oración. Todas construyen sus preguntas según las mismas reglas básicas. Sería mucha casualidad, si no actuara algún principio subyacente de raíz biológica.
La isla de Nueva Guinea es una de las zonas del mundo de mayor densidad lingüística. Fue necesario inaugurar un nuevo tronco lingüístico para su clasificación: la familia Papua, que incluye idiomas como el asmato, el marindo, el chimbú, etc. Pese a todo, esas lenguas responden exactamente a los mismos principios básicos de las restantes lenguas del mundo. Cuando los lingüistas estudian las lenguas a lo ancho y largo del planeta, una realidad se impone: no hay lenguas más sofisticadas o primitivas que otras. Hay pueblos menos avanzados desde el punto de vista tecnológico, pero sus lenguas no son primitivas. Puede haber comunidades en la Edad de Piedra, pero las lenguas que hablan no son de la Edad de Piedra; al contrario, son tan complejas como cualquier otra. Es la constatación de un fenómeno real.
Lenguas criollas
¿Es posible que una lengua emerja de la nada, en el plazo de una generación? En cierto modo es lo que sucede con las lenguas criollas. Han surgido, sobre todo, en un contexto de economía esclavista, donde se juntan cientos de esclavos o trabajadores de orígenes geográficos muy dispares y forzados a convivir durante años. Lo que suele pasar en estos casos es que, al principio, se crea espontáneamente una jerga rudimentaria, un lenguaje macarrónico (pidgin) que permite comunicar mensajes elementales relacionados con el trabajo o la subsistencia. Lo extraordinario es que la generación siguiente, los hijos de esos esclavos o trabajadores, desarrollan una lengua nueva (criolla) con el mismo grado de complejidad que cualquier otra lengua del mundo.
Se plantea que todas las lenguas criollas, aparecidas en sitios muy diversos y sin conexión entre sí, presentan importantes semejanzas estructurales, lo que sugiere que el elemento que es común a todas ellas podría constituir la base de la adquisición del lenguaje por los niños de cualquier parte del mundo.
Bases genéticas del lenguaje
La demostración definitiva de la fundamentación biológica del lenguaje sería encontrar sus mecanismos genéticos. ¿En qué medida el lenguaje es genético y en qué medida es una conducta aprendida? Hoy en día no hay duda de que el lenguaje es el resultado de ambos factores, aunque algunos investigadores atribuyen un papel principal a la genética, y otros, al ambiente. Por una parte es fundamental el papel de los genes para que se produzca la formación de las áreas cerebrales implicadas en el lenguaje y, por otra, lo es el del ambiente para que esas áreas lleguen a realizar su función. Una alteración en los genes acarrea trastornos lingüísticos, como ocurre en la alteración conocida como trastorno específico del lenguaje o en la dislexia. Pero también alteraciones ambientales pueden impedir el desarrollo normal del lenguaje como ocurre en los casos de niños criados en un ambiente de privación lingüística. La mayoría de los investigadores consideran que deben ser varios los genes implicados en el lenguaje, pero hay uno cuya relación con el lenguaje parece estar fuera de toda duda, hasta el punto de que se lo conoce con el nombre de «gen del lenguaje», que es el FOXP2. Sin embargo, no es un gen específico del lenguaje, ya que lo que hace es regular la expresión de otros muchos genes durante el desarrollo embrionario.
De acuerdo con Stromswold (2001), si los estudios genéticos fallaran en revelar evidencias de la genética del lenguaje, ello podría deberse a dos razones distintas. Una posibilidad es que el lenguaje no sea heredable ni tenga tal base genética. Otra posibilidad es que la hipótesis de la herencia del lenguaje sea correcta, pero que su base genética sea la misma para todos los individuos. El caso paradigmático ha sido el de la familia KE, que llamó la atención de la comunidad científica a raíz de una publicación en los años noventa (Hurst et al., 1990). Desde entonces se han realizado varios estudios en esta familia.
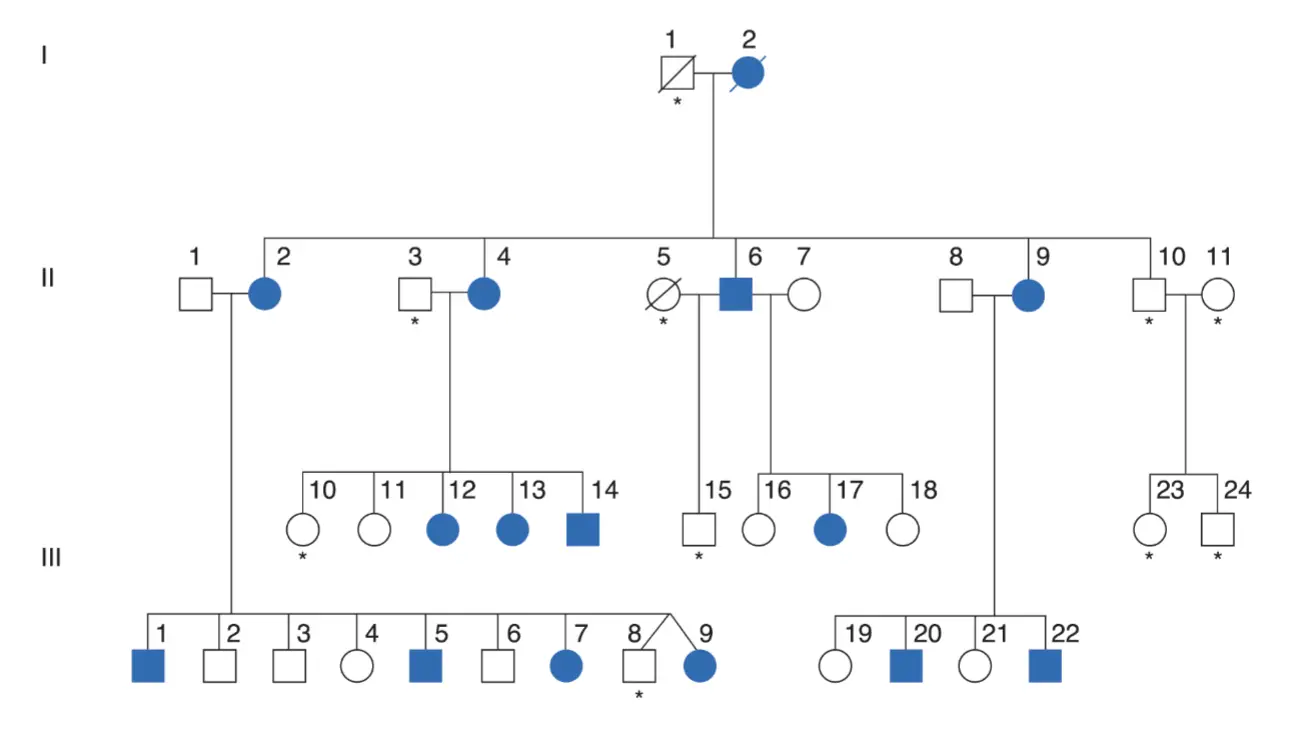
Muchos de los miembros de esta familia presentaban una notable alteración en sus capacidades lingüísticas unida a dispraxia verbal. Este fenotipo es compatible con la acción de un alelo dominante. Estudios moleculares complementarios demostraron que los individuos afectados, y los que no lo estaban, eran portadores de una mutación sin sentido del gen conocido como FOXP2. Otras mutaciones observadas de este gen, concretamente una mutación puntual sin sentido en otra familia diferente, también ocasionara un fenotipo lingüístico similar, no deja duda acerca de la relación entre genotipo y fenotipo en este locus del gen FOXP2. En 2001, un equipo de la Universidad de Oxford dirigido por Anthony Monaco encontró una secuencia de ADN que aparecía alterada (mutada) sistemáticamente en todos los miembros de la familia KE afectados (Lai et al., 2001). El hallazgo se vio reforzado al descubrirse otra persona (CS), que nada tenía que ver con la familia KE, con los mismos síntomas y cuyo ADN también presentaba una mutación semejante (Lai et al., 2000).
La alteración se localizó en el cromosoma 7. La mutación en era mínima, simplemente una base nitrogenada había sido reemplazada por otra (adenina por guanina) en un punto determinado de la secuencia, pero esta sustitución tenía consecuencias en cadena incalculables sobre la síntesis posterior de aminoácidos y proteínas. El nombre del gen donde ocurría la mutación es FOXP2 y pertenece a una familia de genes (genes forkhead) de gran relevancia genética. Así, comenzó a hablarse del FOXP2 como el gen del lenguaje, lo cual todavía es una cuestión abierta, en términos estrictamente científicos.
Como ya señalamos, el papel exacto del gen FOXP2 en relación con el lenguaje es aún controvertido. Su interpretación ha atravesado diversas etapas, dependiendo de la caracterización efectuada del trastorno que provoca. En una primera perspectiva se hablaba de un déficit fundamentalmente lingüístico que afectaba a la adquisición de las reglas morfosintácticas; de hecho, se llegó a llamarlo el «gen de la gramática», concepto que encajaría perfectamente en la teoría de Chomsky y sus seguidores. Posteriormente, se lo consideró más como un gen implicado en la secuenciación compleja de patrones motores articulatorios, patente sobre todo, en la tarea de repetición de seudopalabras (Watkins, Dronkers y Vargha-Khadem, 2002).
Hoy en día el gen FOXP2 ya no se considera el «gen del lenguaje», sino un gen relacionado con habilidades articulatorias y con efectos moduladores sobre otros genes que afectan a habilidades cognitivas (Watkins et al., 2002).
Manual Neuropsicología del Desarrollo
El locus 7q31 del FOXP2, también aparece asociado en algunos estudios con otros cuadros, como el autismo o el síndrome de Gilíes de la Tourette. Los trabajos existentes sobre vinculación entre genes y trastornos del lenguaje escrito apuntan hacia una heterogeneidad genética. En el caso de la dislexia, se han identificado hasta seis locus distintos del genoma verosímilmente asociados con la disfunción lectora. No hay que olvidar que el hipotético enlace entre estructuras genéticas y lenguaje, en caso de existir, probablemente es complejo y no guarda una relación de uno a uno entre genes y funciones. Esto es lo habitual en muchas características hereditarias de naturaleza poligénica, es decir, en las que intervienen varios genes para un rasgo. Nadie sabe en realidad cuántos genes, de haberlos, son necesarios para construir la capacidad lingüística y, lo que es más importante, cómo interaccionan estos genes entre sí.
¿Existe un período crítico para el lenguaje?
Sabemos que existe una relación entre la maduración biológica del individuo y su desarrollo lingüístico, de manera que ciertas etapas no pueden alcanzarse antes de una edad determinada. ¿Existe un período en la vida del ser humano durante el cual está especialmente predispuesto para desarrollar el lenguaje y, una vez pasado, ya no es posible alcanzar un lenguaje normal si, por cualquier circunstancia, se ha visto privado de un entorno lingüístico en su transcurso? El principal defensor de la existencia de un período crítico para el lenguaje ha sido Eric H. Lenneberg en su libro, ya clásico, Fundamentos biológicos del lenguaje (Lenneberg,1967) situaba dicho período entre los 2 años de edad y el comienzo de la adolescencia. Lenneberg se basó fundamentalmente en cinco argumentos, hoy, algunos de estos argumentos han perdido parte de su fuerza:
- Según Lenneberg, los niños con síndrome de Down y que presentan un retraso general en su capacidad mental nunca llegan a alcanzar una competencia lingüística completamente normal, debido a que el desarrollo del lenguaje se detiene al pasar la pubertad, aunque, en teoría, aún quedaría un trecho por recorrer.
- Según Lenneberg, si un niño menor de 2 años sufre una lesión en el hemisferio cerebral encargado del lenguaje (el izquierdo, en la mayoría de las personas), el desarrollo del lenguaje apenas resultaría alterado porque el otro hemisferio se haría cargo de esa función; esto ocurriría incluso en los casos más extremos de pérdida completa del hemisferio izquierdo. La razón de ello residiría en que el período crítico aún no ha comenzado y la plasticidad del sistema nervioso permitiría que el control del lenguaje pasara a depender de otras estructuras neuroanatómicas no afectadas. Estas mismas lesiones suelen producir, sin embargo, secuelas menos reversibles en adolescentes y adultos, porque ya se ha perdido la plasticidad inicial y se ha fijado la especialización funcional de las diversas estructuras neurales.
- La adquisición de una segunda lengua ha constituido otra fuente de argumentación a favor del período crítico. Existen datos que indican que el acento proporcionado por la lengua materna se hace prácticamente inmutable después de la pubertad. Un adulto que aprende un nuevo idioma tiende a conservar su antiguo acento y lo más probable es que su habla nunca llegue a confundirse con la de un nativo, desde el punto de vista fonético. Sin embargo, un niño adquirirá la segunda lengua sin ningún acento extranjero.
- Para Lenneberg, el proceso de especialización hemisférica en la función lingüística se desarrollaría entre los 2 y los 14 años de edad, es decir, el mismo período durante el cual se adquiere y se desarrolla plenamente el lenguaje. De este modo, la capacidad de adquisición lingüística guardaría estrecha relación con el paulatino proceso de especialización hemisférica, gracias al cual, una mitad del cerebro, la izquierda en la mayoría de las personas, se va haciendo cargo de los principales mecanismos neurales del lenguaje.
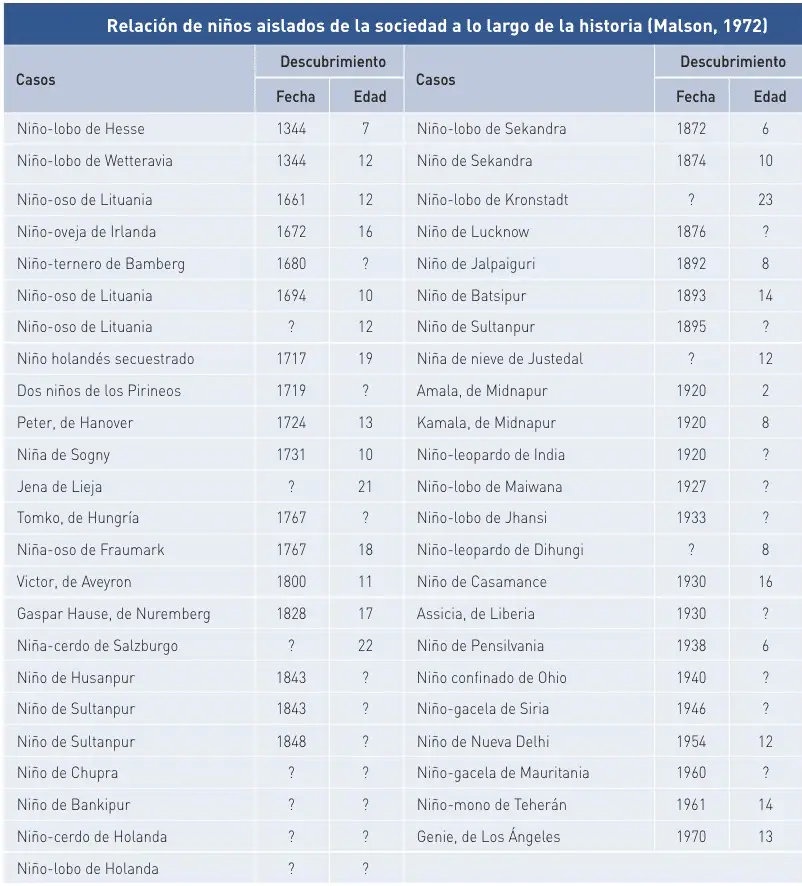
- El quinto argumento se apoya en los individuos que se han visto expuestos al lenguaje tardíamente. Los casos más llamativos son los que se ha dado en llamar «niños salvajes», criados sin la presencia del ser humano, o aislados de toda forma de estimulación lingüística.
Revisión de los argumentos de Lenneberg
Hay autores que entienden que el estancamiento del lenguaje en la pubertad de los niños con discapacidad psíquica se debe, sobre todo, a la falta de estimulación más que al hecho de haber sobrepasado el supuesto período crítico. Por otra parte, algunos estudios más recientes han puesto de relieve que los niños con síndrome de Down presentan un techo preestablecido en su desarrollo y pueden alcanzar este punto a cualquier edad, lo que en muchos casos sucede bastante antes de llegar a la adolescencia (Gleitman, 1984).
Según Aitchison (1992), en lo que a la edad de inicio se refiere, Lenneberg se equivoca al suponer que los niños menores de 2 años no resultan afectados por lesiones en el hemisferio izquierdo. Por el contrario, los bebés que han sufrido una ablación de este hemisferio en su primer año suelen tener problemas importantes de lenguaje. En cuanto a la terminación repentina del período crítico al llegar a la adolescencia, se ha comprobado que a esa edad aún pueden producirse cambios sustanciales en el dominio del lenguaje.
Hay cierto consenso en admitir un subperíodo crítico para un aspecto muy concreto: la fonética de una segunda lengua. Parece que a partir de cierta edad, que algunos autores fijan en torno a los 10-12 años, y otros bastante antes, hay ciertos rasgos de la fonética materna que inevitablemente «contaminan» a la segunda lengua y le confieren un acento extranjero a oídos de los hablantes nativos de ésta. Quien empieza a aprender una lengua a partir de esa edad, probablemente nunca llegará a pasar por un hablante nativo en su pronunciación, por mucho entrenamiento que reciba. Algunos autores, como James Flege, tienden a pensar que la verdadera dificultad del hablante tal vez no resida tanto en la pronunciación, como en la percepción de los sonidos de la segunda lengua: es decir, no hablarían como un nativo porque, en realidad, no percibirían los sonidos de la segunda lengua como un nativo; pero esto es sólo una hipótesis.
Para Lenneberg, la especialización cerebral ocurre entre los 2 y los 14 años de edad, pero, a la luz de lo que sabemos hoy, puede decirse que acontece bastante antes de lo que él supone. Hay incluso bebés menores de 1 año que ya dan muestras de lateralización. En tareas experimentales de escucha dicótica, los niños de 21/2 a 3 años revelan un uso preferente del hemisferio izquierdo en el procesamiento del lenguaje (Kinsbourne y Hiscock, 1987). Según otros autores, el proceso de lateralización ha finalizado en lo esencial hacia los 5 años de edad.
La principal debilidad del argumento de los «niños salvajes» es que no puede descartarse la existencia de un déficit de origen en estos niños.
Niños salvajes
En la mayoría de las ocasiones, la información disponible ha sido muy escasa; a veces, un simple recorte de prensa. Unos pocos casos se han estudiado con más detalle, como en las historias de Victor de Aveyron, Gaspar Hauser de Nuremberg, Amala y Kamala de Midnapur y Genie de Los Angeles. Sólo algunos autores describen las capacidades lingüísticas de los niños al ser descubiertos; en estos casos, todos coinciden en la ausencia de habla y en su incapacidad para entenderla. La mayoría de los intentos de enseñarles a hablar terminaron en fracaso. Se dice que los casos de 1694 (niño-oso de Lituania), 1731 (niña de Sogny) y 1767 (niña-oso de Fraumark) llegaron a hablar un poco, y que Tomko de Hungría, descubierto en 1767, consiguió aprender eslovaco y alemán. También se ha dicho que el niño de Bankipur del siglo pasado aprendió algo del lenguaje de los signos. Pero la información es insegura y los resultados de los casos mejor documentados suelen ser modestos (Crystal, 1994).
El caso de Victor, el niño salvaje de Aveyron, es tal vez el más conocido y considerado como el prototipo de «niño salvaje». Un joven y ambicioso médico llamado Jean-Marc Gaspard Itard fue quien le dio el nombre de Victor y le dedicó un esfuerzo continuado durante varios años. Inventó una gran variedad de juegos y actividades para ayudar a que Victor se socializara y desarrollara alguna forma de lenguaje. Algunas de las técnicas que ensayó se usan hoy en las guarderías infantiles. Todos los intentos de que imitara el lenguaje oral terminaron en un fracaso rotundo, a pesar de que consiguió notables avances en la lectura y la escritura. Para Itard resultaba desconcertante esa disparidad tan marcada en Victor entre la modalidad verbal y la escrita, puesto que jamás consiguió hablar más de una o dos palabras. Se sabía tan poco de la infancia de Victor que nunca se pudo descartar el que el niño fuera mentalmente retrasado o padeciera alguna lesión cerebral congénita.
Mención aparte merecen determinadas historias de privación lingüística por causas sensoriales, como la célebre Helen sordomuda y ciega, o Isabelle, que vivió aislada con su madre sordomuda. Helen Keller fue ciega y sorda desde la infancia a causa de una enfermedad contraída a los 19 meses de vida. A la edad de 7 años, sus padres contrataron a Anne Sullivan Macy, que alcanzó un notable éxito enseñándole el lenguaje a través del sentido del tacto. Helen aprendió a hablar tocando directamente los órganos articuladores (boca, labios, garganta) de Sullivan y otras personas. Aprendió a leer y escribir con el alfabeto Braille y se graduó en la Universidad de Harvard, llegando a ser una brillante escritora y una magnífica profesora al servicio de las personas discapacitadas (Steinberg, 1993).
Isabelle (un seudónimo) estuvo encerrada con su madre muda y no empezó a aprender el lenguaje hasta los 61/2 años de edad. Su madre había sufrido una lesión cerebral a los 2 años de edad y, como resultado de ello, no había desarrollado habla alguna; no sabía leer ni escribir, no recibió educación y se comunicaba con su familia a través de gestos primitivos de su propia creación. Al cumplir los 22 años tuvo a su hija Isabelle. Durante el embarazo y los años siguientes la madre y su hija vivieron confinadas en una habitación, hasta que ambas escaparon y el caso llamó la atención de las autoridades. Helen e Isabelle alcanzaron un nivel de desarrollo lingüístico muy superior al de Victor de Aveyron y Genie de Los Angeles.
Comparándolos, Danny D. Steinberg señala tres factores como principales responsables de la diferencia:
- La edad de comienzo en que ocurrió la privación lingüística.
- La duración de la privación.
- La posibilidad de que existiera desde un principio cierta incapacidad o trauma físico o psíquico en los casos de Victor y Genie.
El caso de Genie de Los Ángeles
En noviembre de 1970 fue encontrada en Los Angeles, en el desván de su casa, una niña de 13 años y medio que había sido criada en el aislamiento más absoluto. El grado de incomunicación y falta de asistencia era equiparable, e incluso superior, al de cualquier «niño salvaje» anterior. Esta niña, conocida como Genie, había vivido en condiciones verdaderamente infrahumanas (Curtiss et. al., 1974). A partir de los 20 meses, Genie fue encerrada en una pequeña habitación […]. Su padre la castigaba si emitía algún sonido. Permanecía la mayor parte del tiempo enganchada con una correa a una sillita de bebé, y cuando no estaba así la introducían en un saco de dormir dentro de una cuna y la tapaban con una tela metálica. Aunque había pasado la pubertad, Genie pesaba sólo 30 kg; no podía masticar alimentos sólidos, no controlaba sus esfínteres y era incapaz de mantenerse erguida. Si hacía ruido, su padre la golpeaba. Nunca le hablaba, aunque en ocasiones le ladraba como si fuera un perro. Su madre, medio ciega y atemorizada por el marido, la visitaba sólo unos instantes diarios y la alimentaba con comidas infantiles; durante este tiempo sólo le dirigía unas pocas palabras, si es que le hablaba. En la casa no había radio ni televisión, por lo que Genie tampoco tuvo ocasión de oír el lenguaje a través de esos medios. Los exámenes médicos parecían indicar que durante los primeros 20 meses había tenido un desarrollo normal hasta que presentó un retraso en la deambulación como consecuencia de una luxación congénita de la cadera. Este hecho hizo que el padre la considerara deficiente mental y la tratara de la forma descrita. Al poco tiempo de que Genie fuera descubierta, su padre se suicidó.
Genie despertó un gran interés (y tal vez ambiciones personales) en el mundo científico norteamericano de los años setenta, por lo que fue sometida a estudio de forma exhaustiva. Hubo en juego, además, importantes dotaciones económicas por parte de los poderosos National Institutes of Health (NIH) estadounidenses. Los tests psicológicos indicaban que su edad mental rondaba los 2 años, con un lenguaje que mostraba muchas de las características de esa edad. Después de unos meses de cuidado, Genie cambió considerablemente. Creció, ganó peso y fuerza y consiguió dar largos paseos. Unos meses después aprendió cientos de palabras sobre nombres de objetos; sentía una enorme curiosidad por los nombres de las cosas que la rodeaban. La persona que estimuló y siguió más de cerca sus progresos lingüísticos fue Susan Curtiss, profesora de lingüística de la (UCLA).
Un año desde su descubrimiento, se realizó una evaluación exhaustiva de los progresos alcanzados por Genie en el lenguaje. Sirva de muestra los siguientes aspectos destacados por Steinberg (1993) en relación con la comprensión oral:
- Comprensión de una amplia variedad de estructuras sintácticas. Por ejemplo, las es-tructuras de negación, al distinguir la frase “Show me the bunny that do es not have a carrot” (Muéstrame el conejito que no tiene una zanahoria) frente a ‘Show me the bunny that has a carrot’ (Muéstrame el conejito que tiene una zanahoria).
- Comprensión de adjetivos simples como ‘big’ (grande) o ‘little’ (pequeño), etc.
- Colocación de los objetos en el espacio en relación con otros objetos, a través de las preposiciones ‘in’ (en), ‘under’ (bajo), ‘next to’ (cerca de), ‘behind’ (detrás), etc.
- Distinción entre singular y plural. Distinción entre las conjunciones ‘and’ (y) y ‘or’ (o).
- Comprensión de pronombres, tiempos verbales y superlativos (‘big’ grande, ‘bigger’ más grande, ‘biggest’ el más grande).
- Preguntas ‘Wh’ (‘who…?’, ‘what…?’, ‘whe-re…?’, etc.) (¿quién?, ¿qué?, ¿dónde?).
Sin embargo, sus mejoras en el lenguaje hablado fueron mucho más lentas. Tardó varios años en alcanzar la fase del lenguaje telegráfico y continuar más allá con expresiones más complejas y maduras. A pesar de que su habla era deficiente y poco gramatical, Genie era capaz de expresar relaciones de significado más complejas, que indicaban que su capacidad intelectual iba por delante de su capacidad lingüística. Sirva de ejemplo el siguiente recuerdo suyo verbalizado: ‘Father take piece wood. Hit. Cry’ (Padre tomar trozo madera. Golpear. Llorar). En los niños normales la diferencia entre el lenguaje productivo y comprensivo decrece con el tiempo y el primero se aproxima al segundo; en Genie tal desfase persistió siempre. En total, Genie fue estudiada durante 5 años; después de ese tiempo sus avances se estancaron. Si se considera su punto de partida, la niña realizó progresos gigantescos, tanto en lo que se refiere a su comportamiento social como lingüístico.
Sin embargo, no fue capaz de adquirir un lenguaje gramatical. No alcanzó aspectos que son dominados por los niños normales antes de los 4 años. Finalmente, un episodio desgraciado, en uno de los muchos hogares adoptivos por los que pasó, la bloqueó emocionalmente contra el empleo del habla. Desde entonces, no volvió a abrir la boca para hablar.
¿Período crítico para Genie? Genie apenas poseía experiencia lingüística antes de la pubertad y, sin embargo, consiguió logros notables después de esa edad, mediante instrucción intensiva y sistematizada. Por otra parte, las condiciones extremas en las que vivió Genie no permiten descartar otras razones como causa de sus dificultades de aprendizaje. No sólo estuvo privada de la oportunidad de oír hablar o de conversar con otras personas, sino que padecía una grave desnutrición, privación social crónica y constantes malos tratos por parte de su padre. No sabemos hasta qué punto sus dificultades con el lenguaje reflejan también las consecuencias de estos abusos y privaciones adicionales.
¿Existen también períodos sensibles en los seres humanos? En ningún caso se pueda hablar de períodos críticos en el desarrollo humano, aunque sí parece claro que determinados aprendizajes pueden ser realizados de forma mucho más sencilla y directa en determinados períodos sensibles. Existen determinadas habilidades, entre la que podemos destacar el reconocimiento de caras o las habilidades de reconocimiento de sonidos que subyacen al desarrollo fonológico, que se verán facilitadas si se tiene la experiencia adecuada antes de que se produzca la «poda» de las conexiones neuronales. Parece claro que la adquisición y desarrollo del lenguaje está sujeto a la existencia de períodos sensibles tanto en la adquisición de los fonemas propios de una lengua determinada, como en lo que se refiere a la adquisición de la gramática, aunque no así en el caso del léxico. Los niños son capaces de aprender rápidamente una segunda lengua convirtiéndose en «maestros» de sus padres, especialmente con respecto a las habilidades fonéticas y gramaticales; a partir de la pubertad, las dificultades del aprendizaje de lenguas extranjeras se incrementan notablemente en la mayoría de los sujetos.
Manual Psicología del Desarrollo I
ÁREAS CEREBRALES IMPLICADAS EN EL LENGUAJE
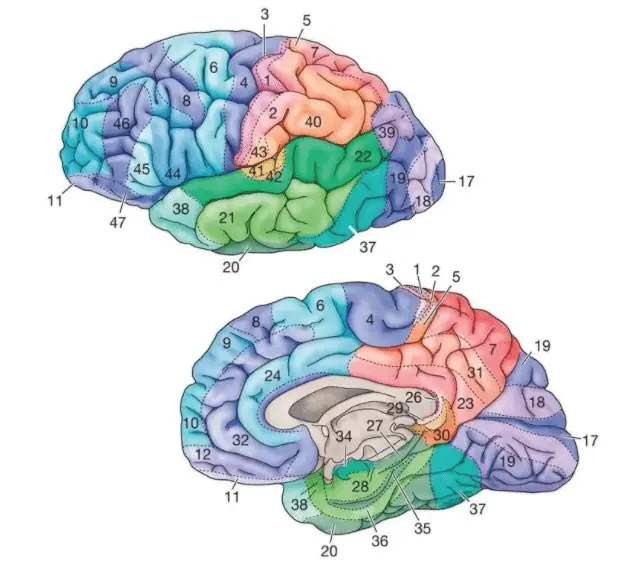
El neurólogo francés Pierre Paul Broca (1824-1880) describió en 1861 el caso, ampliamente citado, del paciente Leborgne. Este enfermo parecía comprender todo lo que se le decía y no mostraba parálisis alguna que afectara a los músculos bucofonatorios, pero sólo podía decir algo parecido a la sílaba «tan». Cuando Leborgne murió, Paul Broca le practicó la autopsia y halló una lesión importante en su cerebro, en el lóbulo frontal del hemisferio izquierdo. Broca presentó sus observaciones a la Sociedad Antropológica y concluyó que esta lesión frontal izquierda, en la zona de la tercera circunvolución, era el origen de la pérdida de la capacidad de hablar del paciente y que era en dicha zona cerebral donde residía la facultad de producir el lenguaje hablado. Un siglo y medio después, las observaciones de Paul Broca se consideran vigentes en líneas generales, y se reconoce la denominada área de Broca como una de las zonas específicamente lingüísticas del cerebro humano, cuya lesión da lugar a la afasia motora o afasia de Broca.
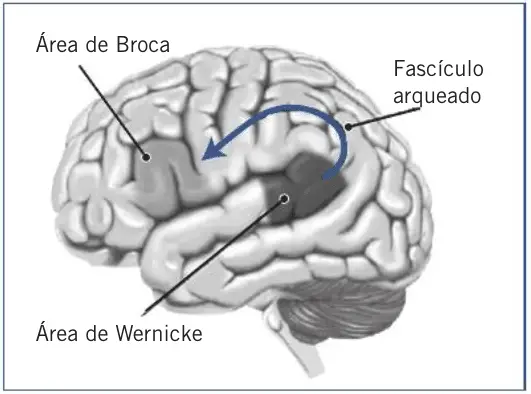
Una década más tarde, el neurólogo alemán de origen polaco Carl Wernicke (1848-1904) describió un nuevo tipo de trastorno de lenguaje. En este caso la lesión cerebral se situaba en el lóbulo temporal, también izquierdo, en una zona próxima al área auditiva, y el paciente presentaba manifestaciones muy distintas a las observadas por Broca. Se trata de la primera descripción de la denominada afasia sensorial, o afasia de Wernicke, en la que existen graves problemas de compresión y el paciente no entiende lo que se le dice. Si la afasia de Broca se caracteriza por un habla lenta y dificultosa, la afasia de Wernicke lo hace por un habla rápida y fluida (afasia fluida), pero vacía de contenido semántico porque el paciente tiene problemas para manejar los significados de las palabras.
Afasia de Broca (motora).
- Expresión no fluida (< 50 palabras/min), habla con dificultad y evidente esfuerzo, articulada pobremente y corta longitud de frase (a menudo una única palabra). Palabras con sentido.
- Tiene problemas en poner en marcha los programas o patrones neuromotores articulatorios de las palabras. El control individual de los músculos está preservado y puede imitar gestos con la lengua, los labios, etc.
- Agramatismo: lenguaje «telegráfico» (omisión de palabras de función). Uso preferente de sustantivos. Dificultad de comprensión gramatical. Uso de estrategias compensadoras, apoyándose en cadenas motoras sobreaprendidas. Por ejemplo, ante la pregunta «¿cuántos años tiene tu hijo?», el hablante puede seguir la serie «uno, dos, tres, cuatro, cinco, seis, siete», debido a la dificultad para pronunciar la respuesta («siete») directamente.
- Notable disprosodia (ritmo y entonación anormales).
- Su comprensión es razonable, pero tiene dificultades para manejar la estructura sintáctica de la frase, por lo que frecuentemente no entiende la relación entre las palabras cuando hay cierta complejidad estructural. P. ej., presenta problemas con las oraciones compuestas, con la voz pasiva o con el orden de las palabras; no entenderá la diferencia entre «Laura insultó a Antonio» y «Laura fue insultada por Antonio».
Afasia de Wernicke (fluida o sensorial).
- Grave defecto de comprensión (del lenguaje ajeno y propio).
- Expresión fluida: muchas palabras, bien articuladas, longitud normal de la frase, prosodia normal. Habla vacía: omisión de palabras de contenido, las palabras carecen de sentido.
- Parafasia (alteraciones y sustituciones de palabras y fonemas). Neologismos. Repetición recurrente de ciertas expresiones.
- Enorme dificultad para comprender el lenguaje y manejar el significado de las palabras de contenido, como los sustantivos, verbos y adjetivos. Los enfermos presentan una gran desorientación para entender no sólo el lenguaje de los demás, sino también el lenguaje propio, con la grave repercusión que esto tiene en su organización del pensamiento.
- En general, se trata de un problema más grave y central que el de la afasia de Broca.
Mientras que los pacientes con afasia de Broca son plenamente conscientes de sus carencias, los de Wernicke no siempre son conscientes del trastorno y a veces incluso niegan su existencia. El neurólogo Ludwig Lichtheim (1845-1928), que trabajó junto a Wernicke, propuso en 1885 una clasificación de las afasias, tanto de las observadas hasta ese momento como de aquellas posibles, aún no descritas. Este sistema clasificatorio, o modelo de Lichtheim, parte de consideraciones neuroanatómicas y plantea un diagrama explicativo que conecta los distintos centros del lenguaje. Este enfoque se ha convertido en un modelo clásico dentro de la neuropsicología, que ha sido desarrollado posteriormente por Norman Geschwind.
El modelo Wernicke-Geschwind
A finales del siglo XIX, Lichtheim y Wernicke plantearon algunos de los principios de la organización del lenguaje en el cerebro humano. Observaron que el área de Broca se halla cerca de la parte de la corteza motora que controla los movimientos de la mandíbula, la lengua, los labios, etc., es decir, los órganos que intervienen en la producción del habla. Por su parte, el área de Wernicke está próxima a la corteza auditiva que recibe la información acústica del entorno y es clave en la compresión de las palabras. Plantearon la hipótesis según la cual ambos centros del lenguaje debían de estar conectados entre sí. En 1970, Norman Geschwind basándose sobre todo en las aportaciones de Wernicke, elaboró un modelo que describe cómo fluye la información entre las distintas zonas cerebrales al usar el lenguaje.
Si se trata de la pronunciación de una palabra escrita, la información se proyecta desde la retina sobre la corteza visual primaria del lóbulo occipital y, de aquí, pasa al área de Wernicke a través la circunvolución angular. En el área de Wernicke probablemente se activa la representación auditiva de la palabra, y la información se transmite al área de Broca a través del fascículo arqueado, y de esta área a la corteza motora, para su pronunciación. Una lesión en el fascículo arqueado causaría un déficit en la repetición del lenguaje, pero no en su comprensión o producción. Es el denominado síndrome de desconexión.
Limitaciones del modelo
En líneas generales, el modelo propuesto por Geschwind sigue siendo válido. Nadie duda del predominio del hemisferio izquierdo en las funciones del lenguaje, ni tampoco del importante papel que desempeñan el área de Broca, el área de Wernicke, la circunvolución angular o el fascículo arqueado. Sin embargo, este modelo es demasiado simple y no recoge ni todas las funciones lingüísticas, que son muchas, ni todas las áreas cerebrales que intervienen en el lenguaje, que también son muchas.
Con las modernas técnicas de neuroimagen se ha podido comprobar el gran número de áreas que se activan cuando se lleva a cabo alguna actividad lingüística y cómo cambian en función de la tarea o de los estímulos. Prácticamente todo el hemisferio izquierdo interviene en el procesamiento del lenguaje: área prefrontal dorsolalteral, área motora suplementaria, áreas temporales superior, media e inferior, zona parietal inferior, circunvoluciones angular y supramarginal. También interviene el hemisferio derecho, especialmente en el procesamiento semántico y en el procesamiento del discurso. Además, participan las estructuras subcorticales, tanto los núcleos grises, principalmente el tálamo y los ganglios básales, como la sustancia blanca, en concreto las conexiones entre esas zonas de la corteza implicadas en el lenguaje (p. ej., el fascículo arqueado) o entre los dos hemisferios (cuerpo calloso) e, incluso, también el cerebelo participa en las funciones lingüísticas.
Dronkers, Redfern y Knight (2000) estudiaron una muestra de más 100 pacientes mediante evaluación neuropsicológica y neuroimagen y comprobaron que algo más del 16% de los clasificados como afásicos de Broca de acuerdo con los resultados neuropsicológicos, no tenían lesión en el área de Broca. Por el contrario, sólo el 50-60% de los pacientes con lesión en el área de Broca presentaban signos de afasia de Broca. En el caso de la afasia de Wernicke las correspondencias eran aun más bajas, puesto que el 35% de los pacientes clasificados como afásicos de Wernicke según las pruebas neuropsicológicas, no tenían lesión en dicha área y el 65% de los pacientes que sí tenían una lesión en esta área no presentaban los síntomas correspondientes a la afasia de Wernicke. En el de las afasias de conducción el desfase era aun mayor, ya que la mayoría de los pacientes no tenían una lesión en el fascículo arqueado sino en la circunvolución temporal superior izquierda y en la parte inferior del lóbulo parietal izquierdo. Por el contrario, lesiones en el fascículo arqueado tenían como resultado graves trastornos de producción, ya que los pacientes sólo eran capaces de repetir sílabas o palabras.
Modelo Friederici
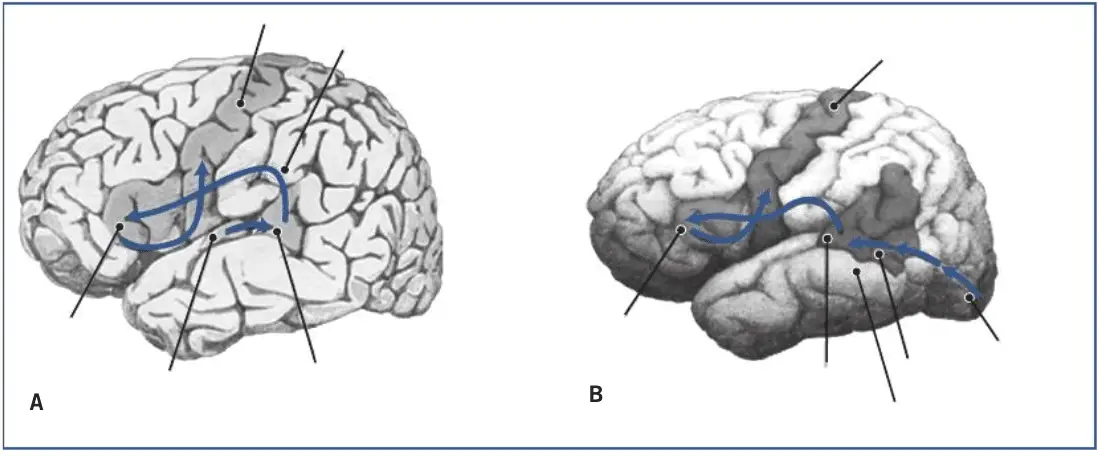
Las modernas técnicas de neuroimagen han podido demostrar que el área de Broca, en realidad, se compone de varias subáreas que realizan funciones lingüísticas diferentes. Angela Friederici y cols. basándose en numerosos estudios de tractografía han propuesto un modelo muy detallado sobre las funciones lingüísticas de las áreas y tractos (Friederici, 2016,2017). El modelo se refiere a los aspectos más nucleares del lenguaje y defiende que el procesamiento sintáctico y semántico tiene lugar en una amplia red cerebral del hemisferio izquierdo, conformada principalmente por cuatro rutas neuronales: dos dorsales y dos ventrales, que conectan diversas regiones frontales con temporales.
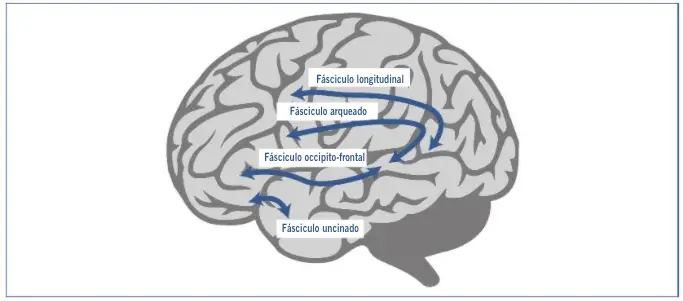
Una de las rutas dorsales, el fascículo longitudinal, conecta regiones de la corteza premotora, área 6 de Brodmann con zonas posteriores de los giros temporales medio y superior, y es esencial en la transferencia y procesamiento de información auditiva-motora; una lesión en ella causa problemas en las tareas de repetición del habla. La otra ruta dorsal, el fascículo arqueado, conecta el área de Broca (área 44 de Brodmann) con las zonas temporales posteriores (área de Wernicke) e interviene fundamentalmente en el procesamiento sintáctico de las oraciones complejas. A su vez, las dos rutas ventrales están involucradas en los procesos semánticos y de estructuras sintácticas básicas. Una de ellas, el fascículo occipito-frontal inferior, conecta las áreas frontales 45 y 47 de Brodmann con los giros temporales medio y superior, y parece estar involucrada esencialmente en el procesamiento semántico. Se ha observado experimentalmente su activación en tareas de categorización semántica de palabras y en juicios sobre la plausibilidad semántica de las oraciones. La otra, el fascículo uncinado, conecta directamente áreas frontales inferiores con áreas temporales anteriores, e interviene en la construcción y el procesamiento de las estructuras sintácticas básicas (Friederici, et al., 2006).
Los experimentos de neuroimagen muestran que esta ruta ventral se activa. por ejemplo, cuando se vulneran las relaciones sintácticas en oraciones básicas formadas por palabras sin sentido. En todo caso, los modelos actuales sobre las bases neurales del lenguaje humano están sometidos a continua revisión, en la medida en que aparecerán nuevas aportaciones empíricas en los próximos años.
Estructuras subcorticales
Las funciones lingüísticas están asociadas fundamentalmente a la corteza cerebral y, en particular, a las áreas de Broca y Wernicke. Sin embargo, en los últimos años se ha ido incrementando la evidencia sobre la importancia de las estructuras subcorticales en el procesamiento del lenguaje. De hecho, la mayoría de los cuadros afásicos corresponden a lesiones cerebrales que no se limitan a un daño cortical, sino que afectan también a estructuras subyacentes cuya participación en el lenguaje todavía es poco conocida. Parece que los núcleos o ganglios basales forman parte de circuitos neurales clave para operaciones de secuenciación, no sólo de conductas motoras, sino también de naturaleza sintáctica (Lieberman, Friedman, y Feldman, 1990). Así, las personas que padecen la enfermedad de Parkinson, en la que hay un daño degenerativo de algunos núcleos basales, como la sustancia negra y el globo pálido, presentan, además de los problemas motores, déficits específicos en el procesamiento sintáctico y gramatical.
Estudios que asocian déficits gramaticales a la enfermedad de Parkinson:
- Illes et al. (1988). Hallaron dificultades similares a las que presentaban los pacientes con corea de Huntington. Las frases producidas por los enfermos de Parkinson a menudo son cortas y tienen una sintaxis simplificada. Al principio se pensó que era una estrategia compensadora frente a los problemas motores propios de la enfermedad, pero trabajos posteriores demostraron déficits sintácticos de comprensión en tareas basadas en respuestas muy sencillas.
- Lieberman, Friedman y Feldman (1990). Encontraron que, los errores de los enfermos de Parkinson tendían a repetirse en el mismo tipo de estructuras de modo regular. La mayor proporción ocurría en frases de ramificación izquierda, del tipo «como estaba lloviendo, la chica jugaba en la casa», o en frases de relativo con ramificación derecha, como «la madre cogió al bebé que estaba llorando».
- Otros estudios independientes también han constatado errores similares en tareas de comprensión y en juicios sobre la gramaticalidad de las frases en enfermos de Parkinson, en particular en el procesamiento de las oraciones pasivas y en la flexión de verbos regulares. Todo esto sugiere que ciertas estructuras y circuitos subcorticales podrían desempeñar un papel importante en algunos aspectos del lenguaje basados en reglas (Teichmann et al., 2005).
Parece que los ganglios basales están involucrados en circuitos neurales esenciales para operaciones de secuenciación. Lo llamativo es que su acción no se limita a la construcción de secuencias motoras, o «sintaxis» motoras, sino que probablemente tienen un papel decisivo en la secuenciación de elementos del lenguaje durante el procesamiento sintáctico. Existen evidencias del papel secuenciador de los ganglios basales en seres humanos y su repercusión sobre el habla y otras funciones. Así, Pickett et al. (1998) presentaron el caso de una mujer con una lesión bilateral, limitada a ciertas estructuras de los ganglios básales, que mostraba una incapacidad para secuenciar los movimientos del habla y, al mismo tiempo, dificultades en la comprensión sintáctica. Sus déficits cognitivos también se focalizaban en operaciones mentales que requerían una correcta secuenciación de elementos. Por otra parte, no parece casual que el único gen (FOXP2) hallado hasta la fecha comprometido en un trastorno probablemente específico del lenguaje interfiera en el correcto desarrollo de los ganglios básales, concretamente del núcleo caudado.
Esta asociación entre circuitos basales y trastornos específicos del lenguaje aparece corroborada en estudios de otra índole. Tallal, Jeringan y Trauner, (1994) observaron una reducción bilateral del núcleo caudado en un grupo de niños con retraso específico del lenguaje. También informaron de una lesión bilateral en la cabeza del núcleo caudado en un niño de 10 años con importantes problemas articulatorios.
ASIMETRÍA HEMISFÉRICA DEL LENGUAJE
El cuerpo humano, al igual que en la mayoría de los animales, es simétrico bilateralmente. Tenemos dos piernas, dos ojos, dos riñones, dos pulmones, etc., y ambos miembros de cada par tienen la misma función. El cerebro también se compone de dos grandes mitades, o hemisferios cerebrales, unidos por un conjunto de fibras nerviosas denominado cuerpo calloso. ¿Conoce la mitad izquierda del cerebro lo que ha aprendido la mitad derecha? Si hablamos de ratas, la respuesta es afirmativa. Esto quiere decir que en el momento del aprendizaje se ha hecho un duplicado de los registros de memoria en ambos hemisferios cerebrales, disponiendo de una redundancia informativa que, tal vez, tiene valor adaptativo y de supervivencia. Sin embargo, es muy probable que esta redundancia sea un lujo que los seres humanos no nos podamos permitir, sobre todo para una función tan sumamente compleja como el lenguaje, que requiere una extensa circuitería neural. A diferencia de lo que ocurre en otros animales, los dos hemisferios del cerebro humano parecen haberse especializado en el tipo de información que procesan. En la mayoría de las personas, el hemisferio izquierdo se ha especializado en el procesamiento de los símbolos lingüísticos. Leask y Crow (2001) analizaron los datos de una muestra de 12.782 individuos. La lateralidad se midió comparando el rendimiento de ambas manos en una tarea que consistía en marcar el máximo número de cuadrados de una cuadrícula durante un minuto. La habilidad verbal se midió con pruebas de vocabulario. Estos autores observaron que las personas con una lateralidad más acusada tendían a ser más precoces en la adquisición de las palabras. Un segundo hallazgo fue que las mujeres, como grupo, además de ser verbalmente más precoces, estaban más lateralizadas que los varones: las diestras eran «más diestras» que los varones diestros. Por otra parte había una proporción menor de zurdas que de zurdos.
Las fuentes de evidencia a favor de la asimetría cerebral o especialización lingüística del hemisferio izquierdo son múltiples y algunas se conocen de antiguo. A continuación pasaremos revista a las principales.
Lesiones
Las dos guerras mundiales proporcionaron abundante material de observación. Los soldados procedentes del frente con metralla alojada en el lado izquierdo del cerebro solían presentar secuelas en el uso del lenguaje, mientras que los que tenían metralla en la parte derecha tendían a presentar el lenguaje intacto o poco afectado. Son conocidos los valiosos trabajos de Alexander Luria .
Hoy en día se dispone de evidencia clínica continua de los cuadros afásicos producidos por lesiones en el hemisferio izquierdo por etiología traumática (sobre todo accidentes de tráfico) o cerebrovascular (embolias, hemorragias cerebrales). El cerebro humano es muy exigente en el consumo del oxígeno aportado por la sangre. Si una arteria cerebral se obstruye por un coágulo o émbolo sanguíneo, se produce un infarto cerebral al interrumpirse la circulación en toda la zona irrigada por ella, con la consiguiente muerte masiva de neuronas por anoxia. Otra causa vascular es la rotura de un vaso sanguíneo que ocasiona una hemorragia intracraneal (conocida también como derrame cerebral). Al no tener ningún espacio libre, ya que el cráneo es indeformable, la sangre extravasada comprime y destruye tejidos cerebrales. Cuando estos accidentes causan anomalías en el lenguaje, la mayoría de ellos se localizan en el hemisferio izquierdo.
Test de Wada
Cuando un neurocirujano tiene que intervenir en el cerebro es muy importante que conozca previamente qué hemisferio de ese individuo concreto se encarga de las funciones lingüísticas, pues podría pertenecer a ese pequeño porcentaje de personas en las que el lenguaje se localiza en el hemisferio derecho o en ambos. En el primer caso, una pequeña imprecisión podría tener consecuencias que no se producirían en el segundo. A tal fin, antes de la operación se suele utilizar el denominado test de Wada. El test de Wada es invasivo, y en los últimos años se tiende cada vez más a utilizar las técnicas de neuroimagen funcional para localizar el hemisferio lingüístico.
Cerebro dividido
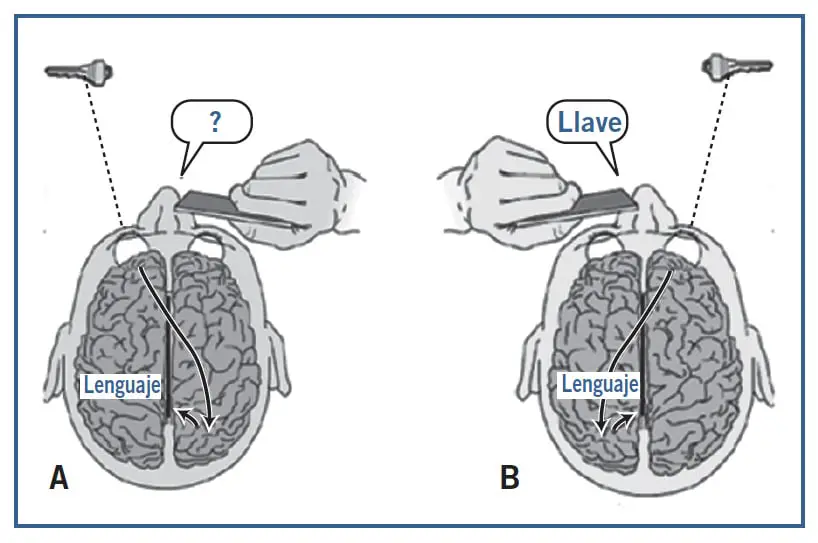
En algunos casos de epilepsias refractarias a otros tratamientos se recurre a una cirugía consistente en la sección del cuerpo calloso para desconectar los hemisferios cerebrales; es lo que se conoce como cerebro dividido o split brain. Por extraño que parezca, las personas con el cerebro dividido pueden llevar una vida aparentemente normal. Sin embargo, algunos experimentos revelan fenómenos muy interesantes en las personas con cerebro dividido. Sperry observó que si un paciente con los ojos cerrados toca un objeto con su mano derecha (p. ej., unas tijeras), puede nombrarlo sin problemas, porque la información táctil viaja directamente al hemisferio izquierdo, donde residen los centros del lenguaje. Pero si lo toca con la mano izquierda, es incapaz de nombrarlo, porque la información se proyecta al hemisferio derecho y desde aquí no puede cruzar al izquierdo para su identificación léxica. No obstante, Sperry constató que el objeto como tal sí es identificado: si luego se le pide que lo localice con la mano izquierda dentro de una caja con otros objetos, el paciente lo hace correctamente. Con la información visual ocurre algo similar. Si se presenta un objeto al hemicampo visual derecho mientras se mantiene la mirada en un punto central, el paciente puede indicar el nombre del objeto, pero si la presentación se hace en el hemicampo visual izquierdo, es incapaz de nombrarlo.
Experimentos conductuales
Múltiples experimentos realizados con personas sanas ponen de manifiesto una asimetría hemisférica respecto a muchas tareas de lenguaje.
Experimentos auditivos
El paradigma auditivo típico es el conocido como audición o escucha dicótica, propuesto por Broadbent en 1954, y que consiste en la presentación simultánea de información distinta a cada oído. Un experimento típico consiste en la presentación simultánea, mediante auriculares, de parejas de sílabas, una distinta a cada oído, por ejemplo, «ba» al oído derecho y «da» al izquierdo, y el participante debe indicar qué ha oído. En otra tarea, ésta de memoria, se presenta la serie completa de parejas de estímulos y se pide al final que el sujeto indique cuáles recuerda.
En general se observa una ventaja del oído derecho en la escucha dicótica de vocales, consonantes, sílabas, palabras y dígitos, mientras que el oído izquierdo (hemisferio derecho) presenta una ventaja en el reconocimiento de melodías musicales, timbres acústicos y otras tareas no lingüísticas, como identificación de emociones o discriminación de intensidades, etc. Hay que señalar que el hemisferio derecho es más eficiente en el procesamiento de la prosodia o entonación del lenguaje (Grimshaw et al., 2003).
Basándose en los resultados obtenidos en la escucha dicótica es posible calcular un índice de asimetría:
índice de asimetría = [(D – I)/(D + I)] x100
- D es la información presentada al oído derecho correctamente indicada.
- I es la información presentada al oído izquierdo correctamente indicada.
El índice de asimetría varía entre -100 (ventaja absoluta del oído izquierdo) y +100 (ventaja absoluta del oído derecho), siendo 0 la igualdad entre ambos oídos. En general, la mayoría de las personas diestras arrojan un índice de asimetría positivo.
Experimentos visuales
El paradigma típico en el dominio visual lo constituyen los experimentos basados en la presentación de estímulos en los hemicampos visuales. Los datos indican que todo el material verbal, como palabras, sílabas, letras, etc., se identifica mejor cuando se presenta al hemicampo visual derecho (hemisferio izquierdo). Sin embargo el hemicampo izquierdo (hemisferio derecho) es superior en el reconocimiento de formas, localización de puntos y otras tareas visuales no verbales ( Springer y Deutsch, 1993). También parece que el hemisferio derecho es más eficiente en el reconocimiento de rostros humanos, lo que concuerda con el hecho de que la prosopagnosia, o trastorno de identificación de las caras familiares, aparece como consecuencia del daño cerebral en determinadas zonas del hemisferio derecho.
Activación del cerebro observada con técnicas de neuroimagen
La quinta fuente de evidencia a favor de la asimetría hemisférica del lenguaje procede de las técnicas de neuroimagen. Gracias a ellas es posible observar qué áreas cerebrales se activan mientras se realiza una tarea. Cuando la tarea es lingüística, la mayoría de las personas presentan mayor activación en el hemisferio izquierdo.
- ¿Por qué es el hemisferio izquierdo el que se encarga principalmente del lenguaje en casi todos los seres humanos? Puede entenderse que esta complejísima función, muy exigente en circuitos neurales, no esté duplicada en ambos hemisferios.
- ¿Por qué en la parte izquierda? Algunos autores consideran que, tal vez, el hemisferio izquierdo posee una ligera ventaja inicial de desarrollo sobre el derecho en casi todos los individuos, lo que explica que también haya muchos más diestros que zurdos. Otros autores consideran factores hormonales como la testosterona, o en diferencias anatómicas entre los hemisferios y en cómo éstos han ido evolucionando en el interior del cráneo, pero parece que no hay nada claro.
- ¿Qué sucede en el resto de los animales? ¿También hay diestros y zurdos? La respuesta es negativa en líneas generales. Un caso aparte, muy interesante por su paralelismo con el habla humana, es el de las aves cantoras. El canto de los pájaros también está funcionalmente lateralizado y depende de centros neurales localizados sobre todo en uno de los dos hemisferios. En el desarrollo de este tipo de asimetría se han buscado implicaciones neurohormonales, porque parece que el control del canto en el canario macho es dependiente de la testosterona. La siringe izquierda, órgano generador del canto, es más pesada y tiene mayor contenido de ciertos componentes bioquímicos que la siringe derecha, pero esta diferencia desaparece en los canarios castrados y, a su vez, puede ser inducida por administración de testosterona en canarios hembra.
METODOLOGÍAS DE INVESTIGACIÓN
En el pasado, la única forma de observar anomalías en el cerebro que pudieran ser causa de trastornos lingüísticos era mediante su estudio post mortem. Los estudios basados en lesiones son valiosos, pero tienen importantes limitaciones. El primer avance significativo se produjo con la aplicación de los rayos X y su posterior implementación gracias a las técnicas de tomografía computarizada (TC), que han permitido pasar de una imagen plana a un modelo tridimensional del cerebro. En los últimos veinte años hemos asistido a una verdadera explosión en el uso de las tecnologías de electrofisiología cerebral y de neuroimagen en la investigación de las funciones del cerebro humano. Permiten investigar la actividad neural que subyace a la realización de los procesos mentales, aunque tienen características distintas.
Potenciales evocados
Esta técnica utiliza pequeños electrodos colocados sobre el cuero cabelludo para detectar la actividad bioeléctrica del cerebro. En realidad, la señal registrada es la del electroencefalograma clásico o EEG, pero tratada mediante procedimientos informáticos para extraer de ella el potencial relacionado con la presentación de un estímulo o ERP. Los ERP disponen de una ventaja frente a las medidas conductuales clásicas: al ser una medida obtenida de forma continua entre la presentación de un estímulo y la emisión de la respuesta, su registro permite evaluar de forma precisa el transcurso temporal de los mecanismos atencionales. Tanto la amplitud como la latencia de los diversos componentes del ERP pueden asociarse a eventos cognitivos específicos. Estos componentes del ERP se designan mediante las letras N o P para indicar si dicho componente tiene un voltaje negativo (N) o positivo (P), respectivamente, seguido de un número que indica el orden serial de ese componente (p. ej., N1 es el primer componente negativo). Dado que los diversos grupos de investigación siguen diferentes convenciones, es necesario observar el eje de ordenadas del ERP para conocer si se han representado los valores positivos arriba y los negativos abajo, o a la inversa.
La presentación de un estímulo, sea visual o auditivo, provoca cambios débiles en el registro eléctrico que normalmente no son visibles directamente porque quedan subsumidos dentro la incesante actividad rítmica u oscilatoria del EEG. A través de procedimientos matemáticos computarizados, que filtran y promedian la señal de múltiples registros correspondientes a estímulos análogos, se eliminan los componentes rítmicos del EEG y se extrae la onda subyacente, mucho más débil, que está relacionada con la presentación del estímulo. Esta onda o señal es propiamente el potencial evocado por el estímulo y es un reflejo, aunque imperfecto, de los procesos cerebrales causados por dicho estímulo.
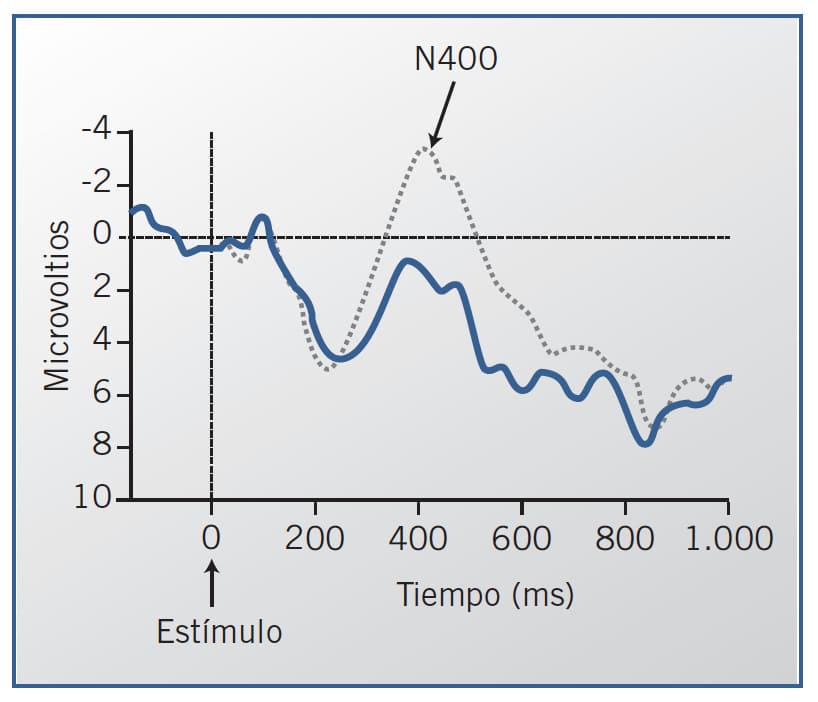
En relación con el lenguaje, un componente muy estudiado, que guarda relación con el pro-cesamiento semántico, es el N400, descubierto por Kutas y Hillard (1980). Consiste en un «valle», o deflexión negativa de la onda cerebral, que surge en torno a los 400 ms tras la administración del estímulo. La situación típica consiste en la presentación de una frase cuya última palabra es anómala desde el punto de vista semántico. Por ejemplo, si se compara la respuesta bioeléctrica ante la frase normal «La pizza estaba demasiado caliente para ser comida» con la respuesta ante la frase anómala «La pizza estaba demasiado caliente para ser bebida», el N400 surgiría alrededor de unos 400 ms tras la presentación de la palabra «bebida». El N400 refleja procesos centrales de comprensión y es independiente de la modalidad sensorial del estímulo, sea visual o auditiva. Está presente incluso ante el lenguaje de los signos para personas sordas. Se puede usar para estudiar el grado de comprensión en personas afásicas.
Magnetoencefalografía
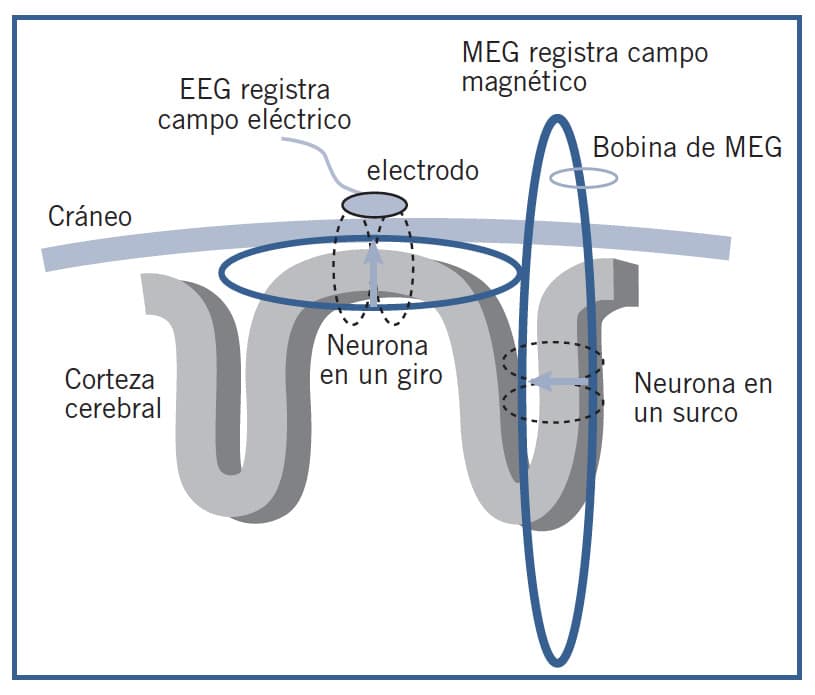
Una técnica cada vez más usada es la magnetoencefalografía (MEG) o registro de los campos magnéticos producidos por la actividad cerebral, a través de unos dispositivos extraordinariamente sensibles. Estos campos magnéticos cerebrales son tan débiles que su registro debe realizarse en cámaras especiales aisladas del campo magnético terrestre. Los microcampos magnéticos se derivan directamente de las corrientes iónicas generadas en las dendritas de las neuronas durante las transmisiones sinápticas. Un único campo es tan débil que no es detectable por sí mismo, pero sí lo es el que surge de varios cientos de millones de neuronas activas a la vez. Las descargas eléctricas de las neuronas registradas mediante el electroencefalograma (EEG) generan campos magnéticos perpendiculares. Dada la anatomía cerebral, el EEG y la MEG proporcionan información sobre la actividad de grupos de neuronas diferentes. Mientras que el EEG registra la actividad bioeléctrica de grupos de neuronas perpendiculares al cráneo, es decir situadas en la superficie de la corteza, la MEG registra los campos magnéticos de grupos de neuronas tangenciales al cráneo, localizadas en los surcos cerebrales.
La MEG tiene buena resolución temporal (inferior a 1 ms) y constituye una buena medida del curso temporal de determinados procesos mentales. Su precisión espacial es mejor que la de los potenciales evocados. En clínica se usa cada vez más para localizar los focos irritativos de pacientes epilépticos o la ubicación exacta de los centros de lenguaje en un individuo antes de una intervención quirúrgica. No obstante, hoy por hoy, la MEG presenta aún dificultades técnicas y metodológicas, sin contar su elevadísimo coste, que hacen que no sea la mejor técnica de exploración en muchas funciones mentales y deba complementarse con otras.
Tomografía por emisión de positrones
La técnica de neuroimagen denominada tomografía por emisión de positrones (PET) se aprovecha del flujo sanguíneo para ofrecer una imagen de la actividad cerebral en un momento determinado. Previamente se inyecta un marcador radiactivo en la sangre que se mezcla con la glucosa contenida en ella. El marcador emite positrones que pueden ser detectados mediante un dispositivo especial, y la mayor densidad de radiación muestra las zonas cerebrales más activas, es decir, las que consumen más glucosa durante el transcurso de un proceso mental determinado. Un ordenador codifica en colores los distintos grados de actividad y ofrece una imagen relativamente detallada de la actividad de cada parte del cerebro. La PET requiere un equipo voluminoso y muy costoso, casi siempre disponible en instituciones hospitalarias para uso clínico. Hay otras técnicas de imagen de detección radiactiva, como la tomografía computarizada por emisión de fotón único o SPECT.
A pesar de sus valiosas posibilidades, la PET es una técnica ciertamente invasiva que requiere la inyección de una sustancia radiactiva en el cuerpo. Aunque la dosis total de radiactividad es relativamente baja, es acumulativa y en los últimos años se han intensificado las críticas hacia su uso.
Resonancia magnética funcional
La resonancia magnética funcional (RMf) se basa en las propiedades magnéticas de la hemoglobina, el componente de la sangre que transporta el oxígeno. La sangre tiene propiedades magnéticas distintas cuando lleva oxígeno (oxihemoglobina), que cuando no lo lleva (desoxihemoglobina). Si la afluencia de sangre hacia una región del cerebro es mayor durante una determinada operación mental, aumenta en esa región la proporción de moléculas de hemoglobina cargadas de oxígeno. De hecho, el ritmo en que el tejido cerebral consume el oxígeno es sobrepasado notablemente por el de su afluencia, por lo que las zonas funcionalmente activas experimentan un estado permanente de sobreoxigenación y, en consecuencia, su tasa de desoxihemoglobina es inferior al de las zonas no activas. Esto tiene consecuencias en la distribución de los campos magnéticos (Howseman y Botwell, 1999; Fiez, 2001).
El estudio de una función o subproceso específico requiere siempre la comparación entre, al menos, dos tipos de imágenes, las obtenidas mientras se realiza la función y las obtenidas en ausencia de dicha función, que sirven de línea base. La diferencia entre ambas indicará las áreas cerebrales que han intervenido durante la función. En algunos experimentos, la línea base se obtiene mientras el sujeto está simplemente en reposo; en otros, el participante realiza una tarea que sirve de comparación con la que se quiere estudiar.
Aunque en la RMf no se inyecta ninguna sustancia radiactiva, como en la técnica PET, no está exenta de limitaciones. En primer lugar, la señal debe registrarse en condiciones de inmovilidad casi absoluta de la cabeza, lo cual excluye o dificulta tareas en las que el sujeto deba dar una respuesta vocal. Se recurre, sobre todo, a tareas encubiertas, de respuesta mental silenciosa, o bien de respuesta motora a través de los dedos sobre una caja especial de respuestas. Las imágenes dependientes de los cambios del flujo sanguíneo tardan segundos en crearse, razón por la cual la resolución temporal es baja y no permite el seguimiento exacto del curso temporal de los procesos. Por otra parte, la relación señal-ruido en una imagen individual es pobre, lo que obliga a promediar muchas imágenes tomadas en las mismas condiciones. El equipo utilizado produce un ruido intenso que resulta problemático para algunos experimentos con estímulos sonoros. Asimismo, el fuerte campo magnético externo impide su aplicación (y sería altamente peligroso) en personas con marcapasos en el corazón o prótesis metálicas. Pese a estos inconvenientes, la RMf ofrece una buena resolución espacial y constituye hoy la técnica de neuroimagen funcional preferida.
Estimulación cerebral no invasiva
Recientemente se ha popularizado un enfoque diferente que intenta demostrar la causalidad cerebro-lenguaje, mediante técnicas de estimulación cerebral «no invasiva», es decir, estimulación magnética o eléctrica realizada externamente a través del cráneo. Algunos de estos métodos tienen una lógica
muy simple; si se inhibe la actividad de una región cerebral mediante estimulación, se creará una «lesión virtual» y deberá producirse una alteración momentánea en la función relacionada con esa región.
Una de estas técnicas es la estimulación magnética transcraneal o TMS (siglas en inglés de Transcranial Magnetic Stimulation). Sobre una zona del cráneo se sitúa una bobina (generalmente en
forma de 8) que envía pulsos magnéticos muy intensos, aunque breves. Los pulsos magnéticos
penetran casi 2 cm en la corteza cerebral y alteran la actividad eléctrica de las neuronas subyacentes.
La estimulación es muy focal, de modo que es importante localizar exactamente el punto del cráneo
que corresponde a la región cerebral de interés. Por ejemplo, para determinar qué punto de la corteza
motora es responsable de la actividad de un músculo concreto de la mano derecha, se aplican pulsos sucesivos sobre la zona central izquierda del cráneo y se registran mediante electrodos los potenciales evocados en el músculo de interés. Se va variando la posición de la bobina centímetro a centímetro hasta que se obtiene un potencial motor visible, que indica la localización precisa de un sector de la corteza motora responsable de la actividad de ese músculo. Algo más complicado es localizar otras áreas funcionales del cerebro que no están asociadas a ninguna actividad motora que se pueda registrar, ya que los individuos difieren considerablemente en la forma del cráneo y, por consiguiente, del cerebro que contiene. En esos casos, se suele obtener una imagen de resonancia magnética del cerebro individual que se incorpora en un sistema de neuronavegación que permite situar la bobina con gran precisión sobre la región cerebral de interés para cada individuo.
La estimulación cerebral no invasiva permite mapear funcionalmente el cerebro, aunque con algunas
limitaciones, ya que sólo se pueden estimular directamente regiones superficiales de la corteza cerebral. Por otra parte, como se trata de un método de intervención, puede utilizarse como herramienta terapéutica.
AUTOEVALUACIÓN
REFERENCIAS
- Cuetos Vega, González Álvarez, Vega, and Vega, Manuel De. Psicología Del Lenguaje. 2ª Edición. ed. Madrid: Editorial Médica Panamericana, 2020.
- Dailymotion
- YouTube